¶ 从神经活动到计算:用于数字分类中模式识别的生物储备池
卢多维科·伊安内洛*¹、卢卡·恰姆皮*¹、法布里奇奥·托内利³、加布里埃莱·拉加尼¹、卢乔·玛丽亚·卡尔卡尼莱²、费德里科·克雷米西、安杰洛·迪加尔博²、朱塞佩·阿马托¹
1 意大利ISTI-CNR
2 意大利IBF-CNR 意大利Bio@SNS
*ludovico.iannello@isti.cnr.it *luca.ciampi@isti.cnr.it
¶ 摘要
本文提出一种基于生物原理的储备池计算方法,其核心是以培养的生物神经元网络作为储备池基质。该系统被称为生物储备池计算,利用活体神经元的自发与诱发活动替(RC)代了人工循环单元。多电极阵列实现了多位点的同步刺激与信号读取:输入通过部分电极传递,其余电极则捕获相应的神经响应,从而将输入模式映射到高维生物特征空间中。我们通过自定义数据集上的手写数字分类案例对该系统进行评估:输入图像经编码后通过电刺激传递至生物储备池,相应的神经活动被用于训练简单的线性分类器。为全面评估生物系统的性能,本文还纳入了与完成相同任务的标准人工储备池的对比实验。结果表明,生物储备池能有效支持分类任务,彰显了其作为可行且可解释的计算基质的潜力。我们认为这项工作为将生物原理融入机器学习提供了更广阔的探索路径,并通过研究活体神经系统如何为设计高效且生物可信的模型提供启示,与受人类视觉启发的计算目标相契合。
¶ 1. 引言
储层计算(RC)[37]是一种通过高维系统动力学转换输入数据的机器学习框架。这种转换通常会产生更易于通过简单分类器分离的表示形式,使得RC成为分类和回归任务(如时间序列分析[3]和语音识别[51])中实用高效的方法。其有效性已在多种实现中得到验证。最著名的RC模型之一是回声状态网络(ESN)[15, 23],它通过大量随机连接的递归单元将输入转换为高维非线性表示,从而促进下游学习。另一个相关范式是液态状态机(LSM)[38,52],它采用脉冲神经元来捕捉丰富的时间动态特性。
在本工作中,我们提出了一种储层计算(RC)范式的实例化方法,其储层并非通过模拟实现,而是通过培养神经元组成的活体网络物理构建。我们将这种方法称为生物储层计算(BRC)。与传统依赖人工单元的RC系统不同,我们的方法利用生物神经培养体系的内在动力学,将输入刺激映射到高维特征空间中。这一架构为探索真实神经组织的计算潜力提供了独特契机,同时也有助于推动将生物学原理整合进机器学习的更广泛目标。事实上,这种基于生物学的方法不仅为计算提供了天然基底,也与神经科学证据相一致——这些证据表明瞬时神经动力学在信息处理中起关键作用[12]。此外,通过将部分计算任务卸载到物理系统中,BRC可能在能效方面具备优势,而这正是现代人工智能系统日益关注的重要议题[2,24]。
具体而言,BRC系统构建于高密度多电极阵列(MEA)[4]之上,该系统既能进行电刺激,又能实现神经活动的高分辨率记录。神经元通过成熟的分化方案[7,8]从干细胞中培育获得,成熟后会形成自发性活跃网络。输入模式被编码为空间分布的刺激序列,并通过MEA传递至培养神经元。由此产生的神经响应被记录并用于构建特征向量,以供后续分类任务使用。为评估BRC系统生成判别性特征表征的能力,我们开展了一项聚焦于单一且具有挑战性的模式识别任务的实验研究:对十种类似数字的空间输入模式(从0到9)进行分类。每种模式由多电极阵列上不同空间构型的电刺激位点定义。记录诱发神经响应并将其作为简单读取层的特征,以对刺激类别进行分类。
尽管生物反应因噪声、自发活动以及跨刺激会话和生物学重复的差异而存在固有变异性,我们的系统仍实现了令人鼓舞的准确度水平。这些结果表明,即使不考虑可塑性或学习机制,培养的神经元网络也能作为有效的储备池,将静态空间输入转化为适用于下游分类任务的丰富高维表征。本论文结构如下:第2节综述关于储备池计算与生物计算的现有文献;第3节详述实验方法,包括MEA接口与刺激协议;第4节展示并分析我们的实验设置与结果;最后,第5节总结研究并展望未来方向。
¶ 2. 相关工作
深度学习(DL)研究的核心议题之一涉及现有计算框架的生物合理性。大量研究批判性地考察了当代神经架构在多大程度上反映了生物神经系统的结构与功能复杂性[18,39,41,48]。鉴于此,学界日益兴起开发受生物学启发的替代方案,旨在拓展机器学习与认知建模的能力边界[34,35]。该方向的努力催生了大量试图弥合人工智能与生物智能鸿沟的模型,这些模型或通过实现更具生物学基础的神经计算[11,14,30,47],或通过改进底层的突触与学习机制[9,21,25,28,29,31-33,36]来达成目标。
在这些受生物学启发的范式中,储层计算(RC)[37, 44] 因其模拟复杂神经动力学的能力而受到广泛关注。RC框架内两个重要的子类别是回声状态网络(ESN)[15,23] 和液态状态机(LSM)[38,52],它们在计算机制和生物学合理性上有所不同。LSM通常采用脉冲神经元模型[1,16]来产生丰富的时序动态,而ESN则依赖大规模随机连接的连续值单元构成递归网络,将输入信号嵌入到有利于下游分类任务的高维空间中。尽管递归架构是随机初始化的,但会采用特定策略确保网络运行在稳定且具有计算效用的动态区间内 。为提高ESN框架的生物合理性,已提出多项改进方案。例如,引入突触可塑性机制(如脉冲时序依赖可塑性STDP [16,46])的模型旨在更贴近地模拟生物学习过程[50]。近期研究中,门控机制的引入增强了非线性循环系统中长期依赖关系的记忆能力[43]。这些受储备池计算启发的技术在语音识别[52]、持续学习[10]等多个应用领域均展现出良好效果。
基于此基础,本研究通过引入生物神经网络作为计算储备池,对储备池计算(RC)范式进行了扩展,即构建了一种生物储备池计算(BRC)体系,其中培养的神经元群体作为计算的动态基底。尽管先前研究已探索利用多电极阵列(MEA)设备与生物神经元交互[13,17,22,26,40,42,45],但仅有少数工作研究了将生物神经网络用作计算储备池的潜力[6,19]。值得注意的是,Cai等人[6]采用脑类器官作为生物储备池,通过利用这类三维生物结构固有的丰富时序动力学特性,展示了其在语音识别方面的能力。他们的方法依托时间依赖的神经活动模式来编码和处理输入序列。与之相反,本研究聚焦于向二维培养生物神经网络施加空间分布式刺激模式,而非三维类器官。具体而言,我们通过微电极阵列(MEA)施加空间图案化电刺激,不依赖时序序列,而是以静态模式识别为目标。这代表了一种独特的范式,强调输入模式的空间编码而非其时间演化。类似地,我们的前期工作[19]探索了使用有限空间图案和网络配置刺激二维培养生物神经网络的可行性。然而,[19]主要是探索性的,涉及少量刺激和应用于单一生物网络的简单分类任务。通过系统研究更广泛的空间分布式刺激模式,并在三个独立生物重复样本(BRs)中评估其效应,本研究成为首次全面评估在储备池计算(RC)框架内使用培养生物神经网络作为静态模式识别功能储备池可行性的重要尝试。该方法不仅 bridging 了生物神经基底与机器学习架构,还为利用生物网络的内在特性——如能量效率和复杂非线性动力学——推进神经形态计算开辟了新途径。
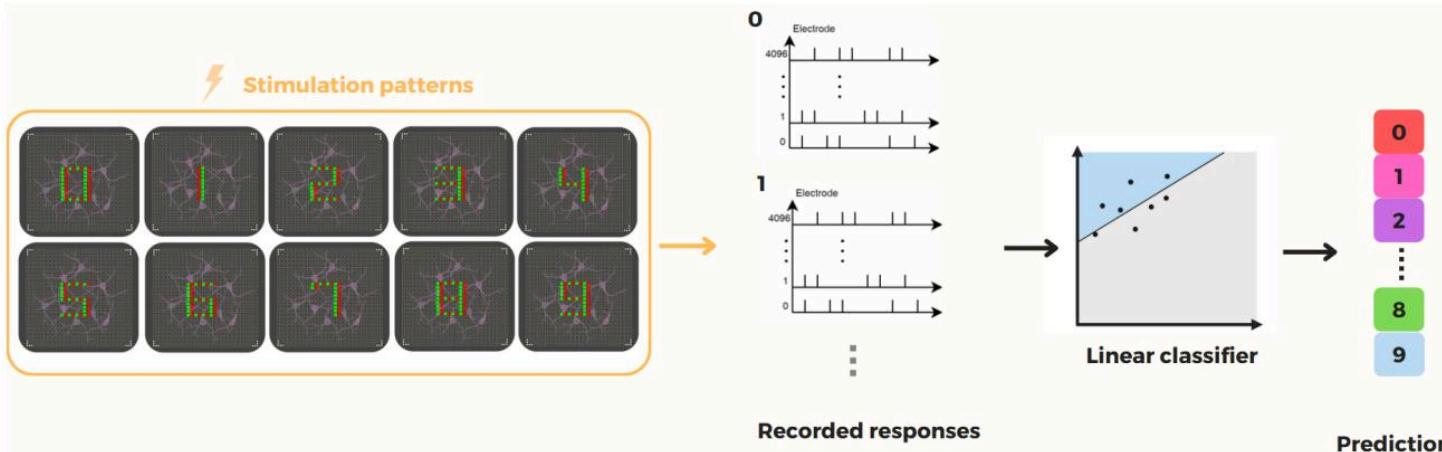
图1. 我们生物储备计算(BRC)范式的框架。在该方法中,多电极阵列(MEA)作为双向接口,既能对培养的生物神经网络进行刺激,又能从中记录信号。离散输入通过选择性激活MEA特定电极子集进行编码,这些电极向网络传递定向刺激。诱发脉冲响应通过另一组电极捕获,并转化为高维向量,在潜在计算空间中编码输入信息。由于生物储备池固有的复杂性和丰富动态特性,这种转换本质上具有非线性特征。随后,训练线性分类器从对应的潜在表征中推断原始输入的类别。

图2. 培养生物网络在MEA装置上的排布示意图。每个方格代表单个MEA电极。输入模式通过将模式元素分配至特定电极,被映射到MEA上。根据相应的输入强度输送电脉冲,并通过其余电极记录网络诱发活动。由此产生的峰值响应用于在生物储备池特征空间中构建输入的高维表示。图中神经元比例未按实际尺度绘制,已调整为便于观察的尺寸。
¶ 3. 生物储备池计算
生物神经元的培养网络可视为一个由随机互连处理元件组成的复杂系统,能够产生丰富的非线性动态行为。当与多电极阵列(MEA)——一种配备电刺激和电生理记录功能的双向平台(见图2)——相连接时,这种神经基质可被功能性利用,以接收结构化输入刺激并产生可测量的响应。
在我们的储备池计算范式中,我们定义了单个输入样本与MEA电极特定子集之间的直接对应关系。例如,当图像被用作输入时,其尺寸为H×W的像素网格会被投影到电极阵列的一个H×W子集上。每个像素都被唯一地映射到一个指定的电极,而像素强度则会调节传递给底层神经元的刺激参数。在刺激阶段之后,我们通过MEA上剩余的电极收集诱发的尖峰活动。这些记录下来的尖峰序列随后被聚合为一个高维特征向量,有效地将输入嵌入到由生物储备池所定义的潜在空间中。
在这种配置下,培养的神经网络充当生物实例化的特征提取器,将结构化输入转化为复杂的、非线性的高维表征[8,42]。
为了定量评估这些生物衍生表征的质量和区分能力,我们采用单层感知器作为下游分类器。完整的计算流程——包括刺激协议和分类框架——如图1所示。以下各节详细介绍了在我们的实验设置中,感知器的刺激、特征提取、训练和性能评估程序。
¶ 3.1. 刺激方案
与生物神经网络接口涉及多项技术挑战,尤其在于刺激参数的校准,例如脉冲幅度、频率和持续时间。必须对这些参数进行精细调节,以确保刺激能引发稳定且强烈的神经响应,同时保持硬件完整性。在本研究中,我们系统性地研究和定义了一套优化的刺激方案,旨在有效激活培养的神经网络,并为可靠的电生理记录提供支持。
为了传递刺激,MEA中的特定电极被指定为有效刺激位点。我们采用双极刺激方式,每对电极配置为相反极性,分别作为正极和负极。每个刺激以矩形双相波形输出,具有设定的振幅A(单位为 )以及分别对应正负成分的相位持续时间 和 (单位微秒)。这种方法增强了正负电极间的电流平衡,有助于维持电极的长期稳定性和完整性。由于MEA将电流分配至多组电极对,振幅需以每对为单位进行解读。在确定这些刺激参数时,需特别注重信号效能与硬件保护之间的平衡:刺激强度必须足够可靠地诱发μ一个 δ尖峰活动,但又不能因强度过高或频率过快导致电极退化——这在涉及高电流或高频率配置的初期实验中已被观察到存在风险。
为了减轻由于顺序效应或时间依赖性可能导致的网络响应偏差,所有刺激模式都以随机顺序呈现给神经元培养物。连续输入之间采用固定的刺激间隔T=10s。这种延迟使网络能够在进行下一次刺激之前恢复到基线或静息状态,从而确保诱发反应的独立性并减少遗留效应[42]。
为在3Brain高密度多电极阵列(HD-MEA)平台上实现此协议,我们基于制造商提供的官方API开发了定制Python脚本。该自定义脚本能够精确控制刺激序列,包括输入顺序的随机化、脉冲发放时序以及特定电极子集的选择,从而确保实验流程的可复现性与灵活性。
¶ 3.2. 生物网络读出
每次刺激呈现时,通过MEA持续记录神经活动,时间窗口为刺激开始前2秒至开始后2秒。为了从原始细胞外信号中实时提取锋电位活动,我们采用了一种基于双阈值策略的锋电位检测算法,该算法能够在线识别锋电位发生的时间。
该算法的运行依赖三个参数:滑动时间窗口 、低检测阈值 ,以及用于最终脉冲识别的高检测阈值 。初始阶段,针对每个记录通道,算法会在时长为 的局部窗口内扫描信号,以识别出超过 的候选峰值——其中 代表完整记录信号的标准差。
为优化检测效果,所有跨越低阈值的信号段会被暂时剔除,并基于剩余数据计算新的标准差 。最终的脉冲时刻则被判定为超过更新后高阈值的局部最大值,该高阈值定义为 :
现将 定义为刺激的起始时刻。对于每个电极 ,我们计算刺激后时长为 的时间窗口内的脉冲发放活动 ——该活动定义为观测到的脉冲数量:
其中, 为二元变量:若在时刻 检测到脉冲,则其值为 1,否则为 0。时间会根据采集系统的采样率进行离散化,在本实验配置中,采样率设为可用的最大频率 。我们将具有恰当定义的时间窗口 的 作为刺激的读出值。最终得到一个4096维特征向量,该向量将输入模式编码至生物储备池的隐空间中。为评估实际网络将信息传播至刺激位点之外的能力,我们从该特征向量中剔除受刺激电极周围的方形区域对应的部分。
¶ 3.3. 分类器训练与测试阶段
单层感知器分类器的训练阶段始于选择一组输入模式,每个模式映射到MEA电极的预定子集进行刺激。对于每个输入样本,通过指定电极施加电脉冲并重复多次,以生成适合统计分析的反应分布。这种重复至关重要,因为生物网络表现出自发活动[49],并且在刺激时刻受到固有噪声和不同动态状态的影响。因此,同一输入模式会在多次试N = 二十验中引发一系列不同的反应。所有特征向量收集完毕后,训练一个线性分类器将每个高维表征与其对应的输入标签关联起来。对于每个类别,记录的反应被随机打乱并分割成互不重叠的训练集和测试集。训练采用通过随机梯度下降法(SGD)优化的单层感知器(SLP)进行[5],目标是最小化交叉熵损失[27]。未使用验证集,因为未采用早停准则;这一选择允许在训练和评估中充分利用现有数据。模型共训练1000个周期。
为评估训练模型的泛化性能,我们采用五折交叉验证策略。在每一折中,数据集被随机划分为五个同等大小的子集。其中四个子集用于训练线性分类器,而剩余一个子集则保留用于测试。该过程重复五次,确保每个子集恰好充当一次测试集。分类器在各折训练结束时达到的状态保持不变,测试期间不进行额外的更新或微调。
由于测试流程复现了训练设置,测试特征的统计特性与训练特征高度吻合,从而确保评估结果具有一致性和无偏性。对于每个测试样本,其潜在表征被输入已训练的分类器,所得预测结果将与对应的真实标签进行比较。系统性能通过分类准确率进行量化,该指标定义为正确预测样本数与测试样本总数的比值。这一指标能够有效衡量该系统作为基于生物学原理的储备池计算架构的性能表现。
¶ 4. 实验评估
本节介绍了用于评估我们所提出的生物储备池计算(BRC)系统的实验装置,并讨论了利用自定义数字数据集得到的实验结果。具体而言,我们通过同一神经培养物的三次生物学重复实验,评估了生物储备池生成的隐表示的分类性能。刺激模式依照电子时钟布局构建,该方法模拟了数字显示屏的显示段——每一对激活的电极对应一个点亮的LED段,共同构成数字的一部分。图3直观展示了所采用的刺激模式,图4则给出了两组记录响应的示例(响应值基于时长为的时间窗口计算得出):
¶ 4.1. 人工储备池对比
为确立参考基准并实现直接对比,我们构建了一个人工储备池(AR)——该储备池基于包含4096个递归单元的网络,单元间的连接稀疏度为10%(即仅10%的潜在单元间连接处于激活状态)。人工储备池以静息态完成初始化,所有单元的激活值均设为0;为保证实验条件的可比性,驱动该储备池的输入刺激与生物实验所用刺激完全一致。
为复现自发神经活动固有的变异性,我们在人工储备池模型中引入了符合生物真实性的输入噪声。该噪声的参数通过经验估计得到:从生物网络记录的自发活动中,随机选取 个时长为 的时间窗口进行分析,再基于这些时间窗口的平均脉冲计数生成合成噪声;合成噪声在输入刺激传入人工储备池之前,被叠加到刺激信号上。每个刺激呈现后,人工储备池的动力学状态会再演化一个时间步;随后参照生物实验中的掩蔽操作,剔除刺激单元周围的方形区域对应的响应,将处理后的结果作为输入,送入单层感知机完成分类任务。
尽管人工储备池模型是极具价值的性能基准,且凭借其工程化的结构有望取得更优的实验结果,但它并不具备真实生物系统的物理特性与能量特性。因此,人工储备池为性能评估提供了上限参考,而生物储备池计算(BRC)系统则为理解基于生物机制的计算模式与高能效信息处理过程提供了关键洞见。
¶ 4.2. 定量结果
为评估生物储备池计算(BRC)系统的性能,首要步骤是确定用于读取网络刺激响应的最优信息时间窗口。为此,我们在多组独立刺激实验中分析了分类准确率,并系统性地改变用于提取神经活动的刺激后时间窗口时长 。
图5呈现的结果,整合了来自 组独立刺激实验的数据,这些实验在不同日期开展,且覆盖了多次生物学重复。结果呈现出明显的下降趋势:当 时,分类准确率始终维持在最高水平。这表明,从不同刺激的可区分性角度而言,包含信息最丰富的响应信号,出现在刺激施加后的最初几毫秒内。
这一结果符合预期:神经响应的初始阶段,主要反映了与受刺激电极直接相连神经元的即时一级激活。从生物学角度来看,5 ms的时间窗口大致对应单次突触传递的时间尺度,能够捕捉到由刺激触发的直接突触后电位。在这一早期时间范围内,信号受到的循环加工、自发活动以及噪声传播的干扰极小,因此对输入模式具有高度特异性。当时间窗口时长 超过该阈值后,记录到的神经活动会越来越多地受到间接响应、网络循环动力学以及自发背景活动的影响。
刺激幅值被设定为每对电极 。所有数字刺激模式均施加于多电极阵列(MEA)的同一区域,该区域的选取依据是其具有最强的自发活动与最优的信号质量。**三类刺激实验(第1天、第2天、第3天)**中各类别的平均分类准确率,以及与人工储备池(AR)在不同噪声实现下平均分类准确率的对比结果,汇总于图6。不同刺激日期及生物学重复实验中,各组刺激实验的平均分类准确率详见表1。
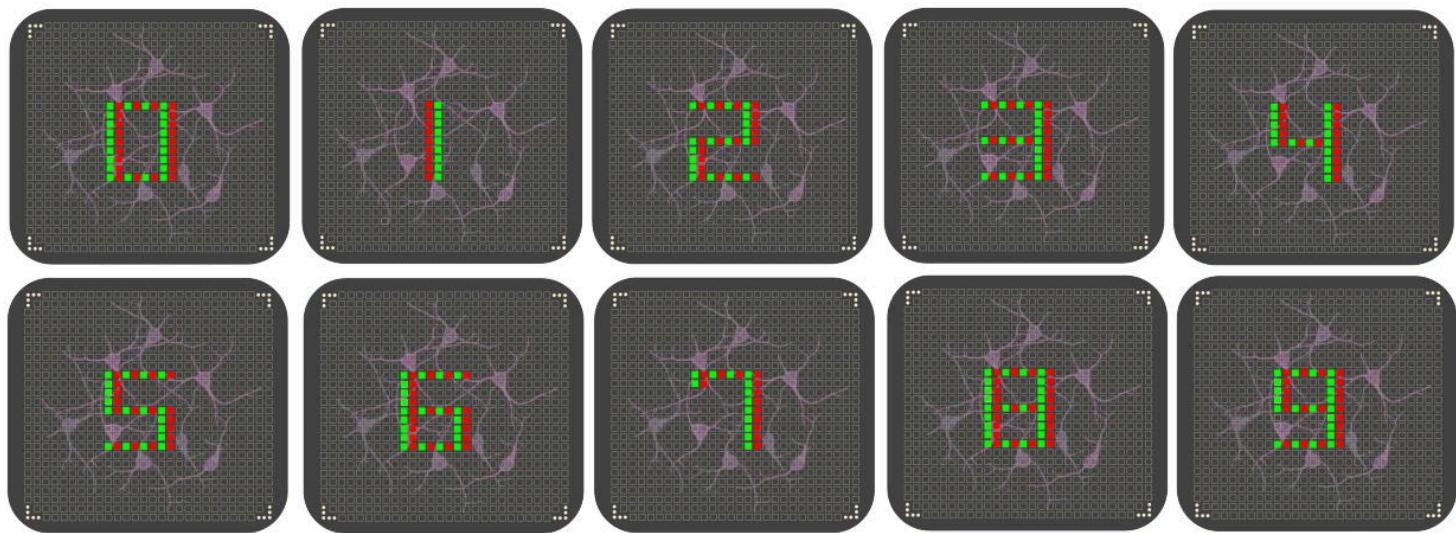
图3.输入模式的可视化表示。数字识别:输入模式表示从0到9的数字。
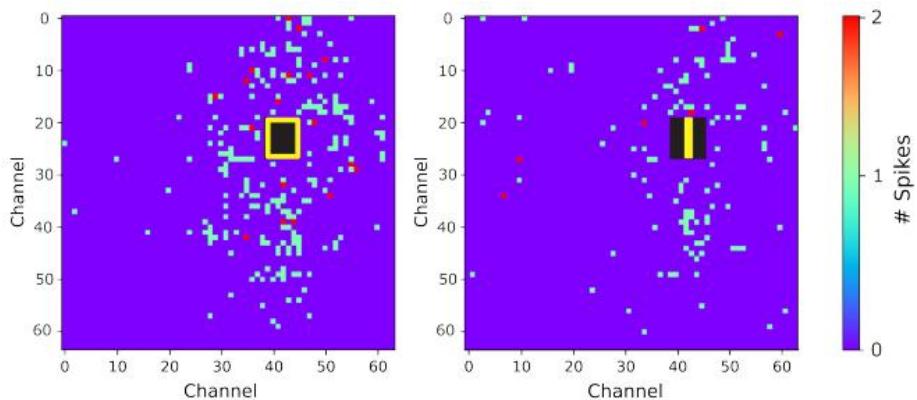
图 4 刺激后神经活动热图:左图展示了输入模式 “0” 刺激后 10 ms 时间窗口内记录到的响应,右图则为输入模式 “1” 对应的响应。热图直观呈现了多电极阵列(MEA)各电极上脉冲发放活动的空间分布特征。
值得关注的是,所有生物储备池均取得了与人工储备池相当的性能水平,平均分类准确率达到约 75%。这一一致性在多组刺激实验以及不同生物学重复中均有体现,凸显了本系统的可靠性与可重复性。尤为重要的是,我们的生物储备池基准充分利用了活体神经基质的固有优势,例如丰富的非线性动力学特性与天然的变异性 —— 这些特性相比传统人工模型,有望提供极具价值的研究洞见与潜在应用优势。
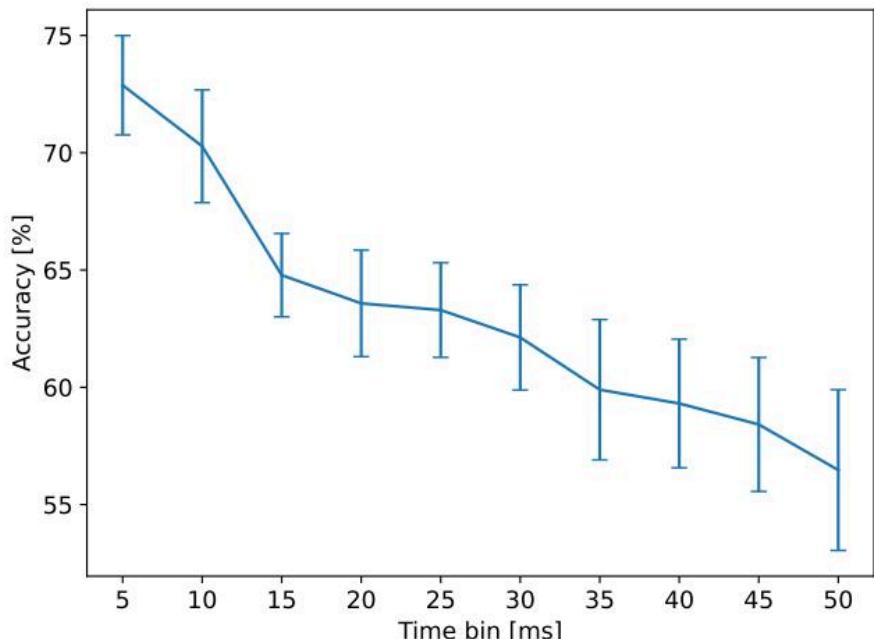
图 5 不同读出窗口下的准确率变化趋势:针对每组刺激实验(n=9),研究人员基于刺激后不同时长窗口(范围为 5 ms 至 50 ms)提取的神经响应数据,计算了分类准确率。该图呈现的准确率结果为均值 ± 均值标准误(SEM)。本项分析揭示了时间整合窗口对储备池读出有效性的影响规律。
¶ 4.3. 准确率随天数的变化
为评估刺激诱发响应随时间变化的稳定性与泛化性,我们采用了如下方案:基于首轮刺激实验的数据训练分类器,再利用后续几天采集的数据对该分类器进行测试。这种跨日测试方法,能够帮助我们探究特定刺激集诱发的神经动力学特性是否在不同日期保持一致,或是会发生自发漂移与重构。
已有充分研究证实,神经培养物的状态会随时间动态演变,其活动模式会自发地在多种状态间转换——从无协调的随机脉冲发放,到高度同步的簇状放电模式[20, 49]。此类自发重构由内在发育进程与突触连接变化驱动,且会对网络响应外部刺激的方式产生显著影响。因此,对于相同的刺激模式,不同日期记录到的诱发响应可能存在明显差异,这一现象反映了网络功能状态的内在转变。
各类别平均分类准确率(均值±标准差)

图6. 每个生物重复样本在各输入类别下的平均分类准确率。每个箱线图展示给定刺激类别的平均分类准确率,该数值通过对每个独立生物重复样本(BR 1、BR 2、BR 3)的多次刺激会话计算得出。准确率采用各会话内的5折交叉验证计算,随后跨三个刺激会话(第1天、第2天、第3天)取平均值。热力图中最后一行显示人工储备池(AR)在不同噪声实现下计算得到的各类别平均分类准确率。
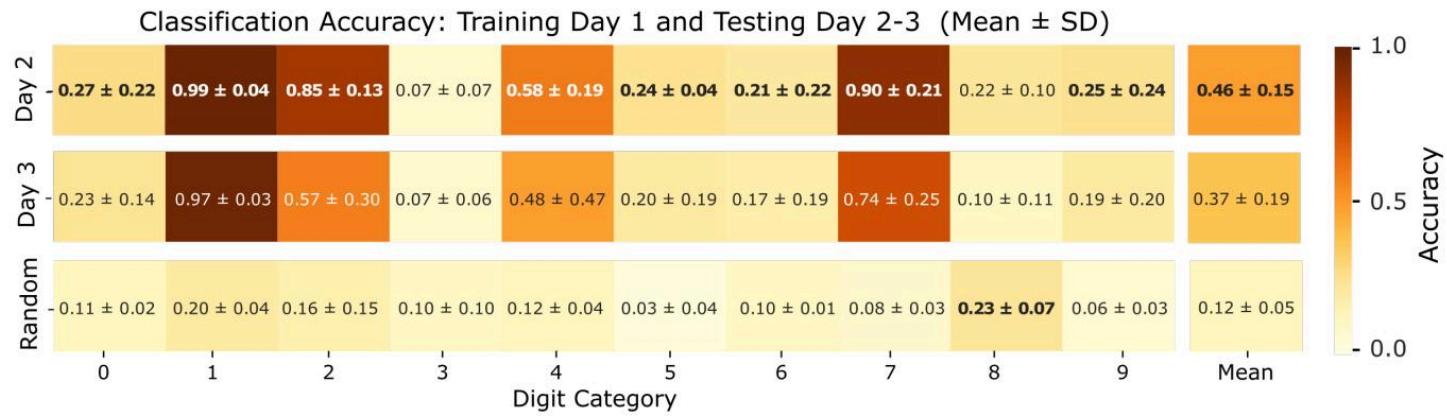
图7. 各输入类别在不同天数上的平均分类准确率。每个箱体代表特定刺激类别的平均分类准确率,其计算方式为:使用第1天的数据训练分类器,并在后续刺激会话(第2天和第3天)上进行测试。准确率为三个独立生物学重复的平均值,每个重复通过五折交叉验证进行评估。误差线表示重复实验间的标准差,反映了生物学和实验因素导致的变异性。最后一行显示的是:通过随机化每个刺激的神经元响应(即保持响应幅度不变但打乱电极记录的尖峰活动空间结构),使用第1天训练的分类器在后续刺激会话上测试获得的各类别平均准确率。
随后,我们利用首轮刺激实验(第1天)中记录到的、神经元对不同刺激模式的全部响应数据,训练了一个线性分类器。接着,我们基于后续独立刺激实验(第2天与第3天)采集的数据,对训练完成的模型进行评估,以此探究生物储备池编码能力的时间稳定性与泛化性。三类生物学重复实验中计算得到的各类别平均分类准确率,汇总于图7。作为额外的对照实验,我们在一个替代数据集上评估了分类器的性能——该数据集的处理方式为:将测试日采集到的每条神经元响应数据,在不同记录通道间进行随机置换。具体操作细节为:针对每种刺激对应的神经元响应,在保留响应信号幅值绝对值的前提下,打乱电极记录到的脉冲发放活动的空间分布结构。不出所料,分类器在该实验条件下无法区分不同的刺激类别,平均准确率仅为 ,这与10分类任务对应的随机猜测水平(准确率)相符。实验结果显示,分类性能随时间推移呈现持续下降趋势:第2天的准确率降至 ,到第3天进一步降至 。尽管这些数值仍高于随机猜测水平(10%),但表明网络随时间维持刺激特异性编码的能力已出现显著衰退。这种性能下降提示,生物储备池内部的神经动力学特性与连接模式会在数天内发生明显演变,这一现象很可能源于神经元培养物固有的自发活动与可塑性效应。
表1. 不同刺激天数与生物重复样本的分类准确率。结果按每次刺激会话(第1天、第2天、第3天)及三个生物重复样本(BR1、BR2、BR3)分别报告。每个数值表示经过5折验证的平均分类准确率与标准偏差。最后一列显示各天跨重复样本的平均性能。
| Stim. day | 生物学重复数(%) | 平均值±标准差(%) | ||
| BR1 | BR2 | BR3 | ||
| Day 1 | 77±5 | 67±9 | 78±6 | 74±6 |
| Day2 | 78±5 | 66±9 | 73±4 | 72±6 |
| Day3 | 72±6 | 68±9 | 67±4 | 69±3 |
有趣的是,两种特定刺激类别——尤其是模式“1”和“7”——在所有实验阶段均持续保持较高的分类准确率。虽然这种稳健性的确切原因尚不明确,但一种可能的解释是:这些模式涉及较少的刺激电极,且与其他刺激的空间重叠极小。这些特性可能导致诱发反应更为清晰、噪声更低,从而更容易被识别。但需要进一步研究来严格评估刺激复杂度、空间干扰与分类稳定性之间的关系。
¶ 5. 结论
本研究提出了一种新型储备池计算(RC)方法,该方法以体外培养的神经元网络作为计算基质。与传统人工储备池不同,我们构建的生物混合系统充分利用了活体神经元的固有动力学特性—— 这些神经元不仅具备极高的能量利用效率,还能表现出丰富的非线性行为。上述特性使生物储备池成为神经形态计算领域极具潜力的替代方案,尤其适用于对低功耗、自适应计算有需求的应用场景。此外,该研究方向还有助于深化学界对生物神经环路信息处理机制的理解,为解析认知功能与神经计算的底层原理提供新的思路。
为验证本方法的有效性,我们基于高密度多电极阵列(HD-MEA)技术,设计并实施了一套定制化的刺激与记录方案。实验聚焦于一项包含 10 个输入类别的数字分类任务(数字 0 至 9),每个数字均对应一种特定的电刺激空间模式。研究人员将记录到的神经元响应数据输入至一个简易的线性读出层进行训练,分类结果证实:生物储备池能够生成高维特征嵌入,足以支持高精度的模式识别任务。上述发现为 “体外培养神经网络可被改造为储备池计算的高效计算模块” 这一观点提供了有力证据,即便是在手写数字分类这类具有一定复杂度的任务中,该系统依然能够稳定发挥作用。
尽管本研究仅采用单一基准数据集评估系统性能,但后续研究将致力于拓展实验范围 —— 通过在更多样、更复杂的输入模式上测试生物储备池计算(BRC)系统,验证本研究结论的普适性。这一拓展性工作将有助于评估生物储备池的可扩展性与泛化能力。
综上,本研究为一类全新的、基于生物机制的储备池计算架构奠定了基础。通过将实验神经科学与机器学习原理有机结合,我们正逐步迈向构建高能效、自适应计算系统的目标,助力缩小人工智能与生物智能之间的鸿沟。
¶ 致谢
本研究得到了Matteo Caleo基金会、比萨高等师范学校(FC)、意大利大学与研究部(MUR)PRIN项目“AICult”(资助编号 #2022M95RC7)(FC)以及欧盟NextGenerationEU资助的PNRR项目“Tuscany Health Ecosystem - THE”(CUP B83C22003930001)的支持。
¶ 参考文献
- Abbott L F, van Vreeswijk C. 脉冲耦合振荡器网络中的异步态[J]. 物理评论E, 1993, 48: 1483-1490.
- Badar A, Varma A, Staniec A, et al. 强调在卷积神经网络中减少研究偏见和碳排放的重要性[A]. AIxIA 2021 - 人工智能进展 - 意大利人工智能协会第20届国际会议论文集[C]. 线上会议: Springer, 2021: 515-531.
- Bianchi F M, Scardapane S, Løkse S, et al. 用于多元时间序列表示与分类的储备池计算方法[J]. IEEE Transactions on Neural Networks and Learning Systems, 2021, 32(5): 2169-2179.
- Bonifazi P, Fromherz P. 用于通过非侵入式接口和模数转换处理实现神经细胞间电子通信的硅芯片[J]. Advanced Materials, 2002, 14(17): 1190-1193.
- Bottou L. 在线学习和随机近似[M]. Cambridge: Cambridge University Press, 1999: 9-42.
- Cai H, Ao Z, Tian C, et al. 用于人工智能的脑类器官储备池计算[J]. Nature Electronics, 2023, 6(12): 1032-1039.
- Chambers S M, Fasano C A, Papapetrou E P, et al. 高效实现人类胚胎干细胞向神经细胞的高度转化 诱导多能干细胞 通过对smad信号的双重抑制[J]. Nature Biotechnology, 2009, 27(3): 275-280.
- Arlotta P, Lancaster M A. 用于体外与体内研究交叉领域中人类神经生物学研究的脑类器官[J]. Nature Neuroscience, 2020, 23(12): 1496-1508.
- Ciampi L, Lagani G, Amato G, et al. 一种生物启发式的生物医学图像分割方法[A]. 计算机视觉 - ECCV 2024研讨会论文集[C]. 米兰: Springer, 2024: 158-171.
- Cossu R, Bacciu D, Carta A, et al. 基于回声状态网络的持续学习[A]. 第29届欧洲人工神经网络、计算智能与机器学习研讨会论文集[C]. 布鲁日(线上): 2021.
- Diehl P U, Cook M. 利用脉冲时间依赖可塑性进行数字识别的无监督学习[J]. Frontiers in Computational Neuroscience, 2015, 9: 99.
- Durstewitz D, Deco G. 皮层网络瞬态动力学的计算意义[J]. European Journal of Neuroscience, 2008, 27(1): 217-227.
- Ferrandez J M, Lorente V, de la Paz F, et al. 生物神经培养物的计算特性研究[J]. 生物医学工程学杂志, 2013, 30(2): 281-286.
- Ferré J, Mamalet F, Thorpe S J. 基于赢者通吃机制的STDP无监督特征学习[J]. Frontiers in Computational Neuroscience, 2018, 12: 24.
- Gallicchio C, Micheli A. 深度回声状态网络(DeepESN):简要综述[EB/OL]. arXiv:1712.04323, 2017.
- Gerstner W, Kistler W M. 脉冲神经元模型:单神经元、群体、可塑性[M]. Cambridge: Cambridge University Press, 2002.
- Gore A, Buonomano D V. 皮层培养物中的时间间隔学习编码于内在网络动力学中[J]. Neuron, 2016, 91(2): 320-327.
- Hassabis D, Kumaran D, Summerfield C, et al. 神经科学启发的人工智能[J]. Neuron, 2017, 95(2): 245-258.
- Iannello L, Ciampi L, Lagani G, et al. 从神经元到计算:用于模式识别的生物储备池计算[EB/OL]. arXiv:2505.03510, 2025.
- Iannello L, Tonelli F, Cremisi F, et al. 神经培养体系中的临界特性见解 深入内嗅-海马网络中的记忆与连接[J]. Chaos, Solitons & Fractals, 2024, 194: 116184.
- Ilin B, Gerstner W, Brea J. 生物可信的深度学习——但浅层网络能走多远?[J]. Neural Networks, 2019, 118: 90-101.
- Isomura T, Kotani K, Jimbo Y. 培养的皮层神经元可根据自由能原理实现盲源分离[J]. PLoS Computational Biology, 2015, 11(12): e1004641.
- Jaeger H. 分析与训练递归神经网络的“回声状态”方法——附勘误说明[R]. Bonn: German National Research Center for Information Technology (GMD), 2001.
- Javed F, He Q, Davidson L E, et al. 大脑及高代谢率器官质量:对去脂体重之外静息能量消耗的贡献[J]. American Journal of Clinical Nutrition, 2010, 91(4): 907-912.
- Journé A, Garcia Rodriguez H, Guo Q, et al. 无反馈的赫布深度学习[A]. 第十一届国际学习表征会议论文集[C]. 基加利: OpenReview.net, 2023.
- Kagan B J, Kitchen A C, Tran N T, et al. 体外神经元在模拟游戏世界具身化时能够学习并表现出感知能力[J]. Neuron, 2022, 110(23): 3952-3969.e8.
- Kline D, Berardi V L. 重新审视用于训练神经网络分类器的平方误差与交叉熵函数[J]. Neural Computing and Applications, 2005, 14(4): 310-318.
- Krotov D, Hopfield J J. 通过隐藏单元竞争进行无监督学习[J]. Proceedings of the National Academy of Sciences, 2019, 116(16): 7723-7731.
- Lagani G, Falchi F, Gennaro C, et al. 使用竞争性赫布学习方法训练卷积神经网络[A]. 机器学习、优化与数据科学——第七届国际会议论文集[C]. 格拉斯米尔: Springer, 2021: 25-40.
- Lagani G, Mazziotti R, Falchi F, et al. 通过精确模拟评估神经元培养物的模式识别性能[A]. 第十届IEEE/EMBS神经工程国际会议论文集[C]. 意大利(线上): IEEE, 2021: 726-729.
- Lagani G, Bacciu D, Gallicchio C, et al. 基于赫布学习的稀缺数据条件下深度特征CBIR[A]. 基于内容的多媒体索引国际会议论文集[C]. 格拉茨: ACM, 2022: 136-141.
- Lagani G, Falchi F, Gennaro C, et al. 使用卷积神经网络比较赫布学习与反向传播学习的性能[J]. Neural Computing and Applications, 2022, 34(8): 6503-6519.
- Lagani G, Falchi F, Gennaro C, et al. Fasthebb:将深度神经网络赫布训练扩展至ImageNet级别[A]. 相似性搜索与应用——第15届国际会议论文集[C]. 博洛尼亚: Springer, 2022: 251-264.
- Lagani G, Falchi F, Gennaro C, et al. 脉冲神经网络与仿生监督深度学习:综述[EB/OL]. arXiv:2307.16235, 2023.
- Lagani G, Falchi F, Gennaro C, et al. 突触可塑性模型与仿生无监督深度学习:综述[EB/OL]. arXiv:2307.16236, 2023.
- Lagani G, Falchi F, Gennaro C, et al. 基于快速赫布规则的可扩展仿生深度神经网络训练[J]. Neurocomputing, 2024, 595: 127867.
- Lukosevicius M, Jaeger H. 储备池计算在递归神经网络训练中的应用[J]. Computer Science Review, 2009, 3(3): 127-149.
- Maass W, Natschläger T, Markram H. 无需稳定状态的实时计算:一种基于扰动的神经计算新框架[J]. Neural Computation, 2002, 14(11): 2531-2560.
- Marblestone A H, Wayne G, Kording K P. 走向深度学习与神经科学的融合[J]. Frontiers in Computational Neuroscience, 2016, 10: 94.
- Pastore V P, Massobrio P, Godjoski A, et al. 基于多电极记录的大规模神经元集群中兴奋-抑制性连接与网络拓扑的识别[J]. PLoS Computational Biology, 2018, 14(8): e1006334.
- Richards B A, Lillicrap T P, Beaudoin P, et al. 面向神经科学的深度学习框架[J]. Nature Neuroscience, 2019, 22(11): 1761-1770.
- Ruaro M E, Bonifazi P, Torre V. 迈向神经计算机:利用神经元培养体进行图像处理与模式识别[J]. IEEE Transactions on Biomedical Engineering, 2005, 52(3): 371-383.
- Scardapane S, et al. Gated Echo State Networks的初步研究[A]. 国际智能系统创新与技术应用会议论文集[C]. 诺维萨德: IEEE, 2020: 1-5.
- Scardapane S, Butcher J B, Bianchi F M, et al. 生物启发式神经网络的优化与应用[J]. Neural Computing and Applications, 2017, 28(12): 2953-2967.
- Shahaf G, Marom S. 大脑皮层神经元网络中的学习机制[J]. The Journal of Neuroscience, 2001, 21(22): 8782-8788.
- Song S, Miller K D, Abbott L F. 通过脉冲时序依赖突触可塑性的竞争性赫布学习[J]. Nature Neuroscience, 2000, 3(9): 919-926.
- Sun Z, Cutsuridis V, Caiafa C F, et al. 大脑模拟与脉冲神经网络[J]. Cognitive Computation, 2023, 15(4): 1103-1105.
- Tenenbaum J. 构建能像人类一样学习与思考的机器[A]. 第十七届自主智能体与多智能体系统国际会议论文集[C]. 斯德哥尔摩: ACM, 2018: 5.
- Tonelli F, Iannello L, Gustincich S, et al. 双抑制mapk/erk和bmp信号诱导内嗅皮层小鼠胚胎干细胞来源的皮层祖细胞中的al样特性[J]. Stem Cell Reports, 2024, 4(7).
- Wang Q, Li P. D-LSM:具备无监督循环储备池调谐的深度液态机[A]. 第23届国际模式识别会议论文集[C]. 坎昆: IEEE, 2016: 2652-2657.
- Yonemura H, Katori Y. 基于储备池计算的动态预测编码实现抗噪声多感官语音识别[J]. Frontiers in Computational Neuroscience, 2024, 18.
- Zhang Y, Li P, Jin Y, et al. 一种具有生物启发学习机制的数字液体状态机及其在语音识别中的应用[J]. IEEE Transactions on Neural Networks and Learning Systems, 2015, 26(11): 2635-2649.